La Francisella philomiragia ha sido aislada de salmón del Atlántico cultivado en balsas-jaulas en el lago Llanquihue, presentando brotes de una enfermedad granulomatosa con altas tasas de morbilidad y una mortalidad acumulada entre el 5% y el 20%. Los aislados bacterianos mostraron un 100% de similitud con F. philomiragia ssp noatunensis o F. piscicida aislados de bacalao en Noruega, un 99% de similitud con Francisella sp. detectado en tilapia en Asia y Centroamérica, y un 99% de similitud con la cepa tipo F. philomiragia mediante análisis filogenético del gen 16s rDNA.
Introducción a la Patología y su Contexto Histórico
A finales de la década de 1980, se detectó en Chile una nueva patología infecciosa cuyo agente causal presentaba morfología rickettsial, denominado inicialmente "unidentified agent". Este agente fue aislado por primera vez en una línea celular de embrión de salmón Chinook CHSE-214 y posteriormente identificado como Piscirickettsia salmonis. La piscirickettsiosis se convirtió en una de las principales causas de mortalidad en salmónidos cultivados en las costas marinas del sur de Chile, generando significativas pérdidas económicas para la industria salmonera.
Sin embargo, a mediados de la década de 1990, se logró aislar un segundo agente con morfología rickettsial en salmón del Atlántico cultivado en el lago Llanquihue. Este agente fue denominado "unidentified agent 2" (U2 o UA-2) para diferenciarlo de P. salmonis. Desde junio de 2006, planteles de salmón del Atlántico cultivados en balsas-jaulas en el lago Llanquihue experimentaron altas tasas de morbilidad y mortalidad, con múltiples brotes simultáneos de una enfermedad crónica de tipo granulomatosa, afectando principalmente el bazo. En todos estos casos, se detectó y aisló un agente bacteriano con morfología rickettsial, sugestivo de U2.
Identificación y Caracterización Molecular de Francisella philomiragia
El análisis filogenético de las secuencias del gen 16s rDNA de los aislados reveló su pertenencia a la familia Francisellaceae, descrita en 2005. Esta familia agrupa bacterias dentro de la subclase gamma proteobacteria y contiene un único género, Francisella. Este género incluye dos especies tipo: F. tularensis, un patógeno humano, y F. philomiragia, aislado de fuentes de agua, ratas de río e infecciones accidentales en humanos. Adicionalmente, existe una gran cantidad y variedad de otras Francisella sp. de afiliación incierta, aisladas de diversas fuentes como garrapatas, el ambiente y una amplia gama de peces.
El análisis filogenético de múltiples genes (16s rDNA, espaciador intergénico ITS, 23s rDNA, rpoB, fopA, groEL, lpnB y sdhA) demostró que los aislados pertenecían al grupo de F. philomiragia, con un 100% de similitud con F. philomiragia ssp noatunensis. Estos aislados formaban parte del mismo grupo junto con otras Francisella aisladas de bacalao en Noruega, y tilapia en Asia y América Central.
Metodología de Investigación
Análisis Bacteriológico, Susceptibilidad Antimicrobiana y Cultivo Celular
Los tejidos de riñón, bazo, hígado, corazón, aletas y branquias de los peces afectados fueron cultivados en diversos medios bacteriológicos bajo condiciones de completa asepsia. La incubación se prolongó por 8 semanas a 18 °C, con monitoreo diario para observar el crecimiento de colonias bacterianas. Las colonias observadas eran redondas, de 1 a 2 mm de diámetro, de consistencia butirosa, convexas, brillantes, de color gris-verdoso y sin pigmento difusible en agar Eugon, con crecimiento visible a los 12 días de incubación a 18 °C.
La tinción Gram de las colonias evidenció estructuras cocoides y cocobacilares Gram negativas, dispuestas individualmente, en pares y en agrupaciones. Las pruebas de oxidasa resultaron negativas y la reacción de catalasa fue débil. En el desafío a diferentes salinidades, los aislados se desarrollaron únicamente entre 0.4% y 1% de NaCl, escasamente a 2%, y no hubo desarrollo a 3% y 6%. En cuanto a la temperatura, los aislados no se desarrollaron a 5 °C, 30 °C y 37 °C a las 8 semanas de incubación. El desarrollo fue lento a 10 °C, 15 °C y 25 °C, y abundante a 18 °C, 20 °C y 22 °C, con desarrollo inicial a los 6 días a 22 °C y 12 días a 18 °C de incubación.
Se determinó la concentración inhibitoria mínima (CIM) para varios antibióticos: florfenicol (1.0 µg/ml), flumequina (0.25 µg/ml), ácido oxolínico (0.25 µg/ml), oxitetraciclina (0.5 µg/ml) y amoxicilina (> 64.0 µg/ml). En los cultivos celulares, se observó efecto citopático (ECP) en OKE-1 a las dos semanas de incubación. En BF-2, se apreció desprendimiento celular de la monocapa a los 12 días, y en SHK-1, ECP y desprendimiento celular a los 20 días. Las líneas celulares CHSE-214 y RTR-2 no mostraron ECP o desprendimiento celular a las ocho semanas, aunque la tinción Gram y la microscopía electrónica revelaron la presencia del agente y cambios ultraestructurales en la morfología celular.
Análisis Histopatológico y Microscopía Electrónica
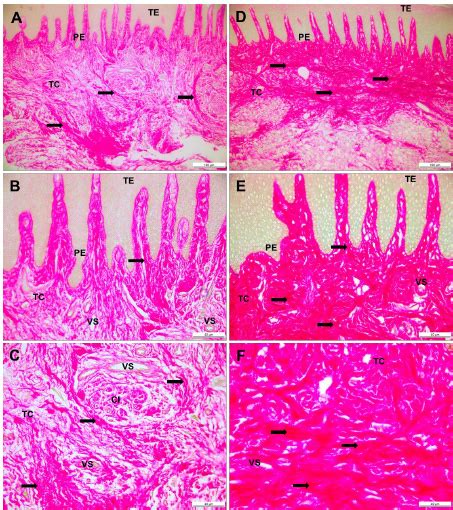
El análisis histopatológico reveló una patología sistémica con degeneración celular y necrosis diseminada en hígado, bazo y riñón. Los bazos mostraron degeneración del tejido conectivo y esplenocitos con núcleos picnóticos y cariorréxicos. Los leucocitos y macrófagos se observaron como células hipertróficas con vacuolas intracitoplasmáticas que contenían agrupaciones de bacterias cocoides basofílicas. En el tejido hematopoyético del bazo se evidenció necrosis extensiva con degeneración y lisis de las membranas nuclear y citoplasmática.
Los riñones presentaron una glomeronefritis crónica y difusa con acumulación de centros de melanomacrófagos en el tejido necrótico. Se observó hepatitis y necrosis ampliamente distribuida, con degeneración hidrópica de los hepatocitos y megalocitosis, y ruptura aislada de los ductos biliares con exudado biliar en el parénquima hepático. La tinción Gram permitió observar bacterias de formas cocoides y cocobacilares Gram negativas, formando agrupaciones intracitoplasmáticas o libres en la matriz extracelular o detritus celular en diversos tejidos.
El análisis por microscopía electrónica mostró agrupaciones de bacterias de forma cocoide y elongada (300 a 1.000 nm) dentro de vacuolas (endosomas o fagosomas) contenidas.
Amplificación y Secuenciamiento Genético
El ADN genómico bacteriano fue extraído utilizando kits comerciales. La amplificación por PCR de los genes se realizó utilizando partidores específicos descritos en la investigación. Las amplificaciones se ejecutaron en un termociclador en un volumen final de 50 µl. El perfil térmico incluyó un paso de desnaturalización inicial, 30 ciclos de desnaturalización, anillamiento y extensión, seguido de una extensión final.
Los productos de PCR se analizaron por electroforesis en gel de agarosa y las bandas correspondientes fueron purificadas. El ADN purificado fue secuenciado utilizando un kit específico en un analizador genético. Las secuencias obtenidas fueron editadas y publicadas en la base de datos pública GenBank con números de acceso específicos para los genes rpoB, groEL, sdhA, fopA, lpnB y 16s ITS 23s rDNA.
Cuadro 1: Conjunto de partidores usados para el secuenciamiento genético
| Gen | Partidor Forward | Partidor Reverse |
|---|---|---|
| 16s rDNA | AGAGTTTGATCCTGGCTCAG | GTGTTAGCAAGCTTCGACGA |
| ITS | TCGTCGCTGCGTGAGTGCA | TTCACGCCCGTTCCGTGC |
| 23s rDNA | AAAGGCCCGGGAAACCCG | GGGTCCGGAAAAGCTCC |
| rpoB | ATGACCCAGTCGGCTGGAT | ATGCCTCGATTCCGCAG |
| fopA | ATGAAAGGTGTCCGTAAT | TCAGCCGTCAGCTGCAG |
| groEL | ATGAAAGGTGTCCGTAAT | TCAGCCGTCAGCTGCAG |
| lpnB | ATGAAAGGTGTCCGTAAT | TCAGCCGTCAGCTGCAG |
| sdhA | ATGAAAGGTGTCCGTAAT | TCAGCCGTCAGCTGCAG |
Análisis Filogenético
El análisis filogenético de los genes se realizó mediante alineamientos múltiples de secuencias de especies bacterianas relacionadas para el gen 16s rDNA. Se utilizó la información de los genomas de Francisella disponibles en GenBank para los genes 16s+ITS+23s rDNA, y un análisis multilocus con los genes constitutivos (housekeeping) rpoB, groEL, sdhA, fopA y lpnB.
Los árboles filogenéticos se construyeron con el criterio de la regla de la mayoría (50%) a partir de 1000 réplicas de bootstrap, utilizando el método de máxima similitud y un proceso heurístico de búsqueda. Se emplearon modelos de sustitución de nucleótidos seleccionados por el software Modeltest para cada análisis.
Resultados
Observaciones de Campo y Necropsia
Los peces afectados presentaron anorexia, natación errática en superficie y letargia. A la necropsia, se observó aumento de tamaño del hígado, riñón y bazo, con múltiples focos blanquecinos en hígado y bazo. La grasa perivisceral estaba presente y la vejiga natatoria se encontraba congestiva. En los casos más severos, se observaron equimosis.
Análisis Bacteriológico, Susceptibilidad Antimicrobiana y Cultivo Celular
Los cultivos bacterianos en diversos medios resultaron negativos a las 8 semanas de incubación, excepto en agar Eugon, donde se observó el desarrollo de colonias características. La tinción Gram reveló cocos y cocobacilos Gram negativos, con pruebas de oxidasa negativas y catalasa débil. El crecimiento se limitó a salinidades bajas (0.4%-1% NaCl) y temperaturas moderadas (18-22 °C).
Las CIMs indicaron sensibilidad a florfenicol, flumequina, ácido oxolínico y oxitetraciclina, pero alta resistencia a amoxicilina. Se observó efecto citopático en diferentes líneas celulares, indicando la patogenicidad del agente aislado.
Análisis Histopatológico y Microscopía Electrónica
La histopatología confirmó una patología sistémica con degeneración y necrosis en órganos vitales. La presencia de bacterias Gram negativas se evidenció mediante tinción Gram y microscopía electrónica, localizadas intracelularmente y en el entorno celular.
El Uso de Antibióticos en la Salmonicultura Chilena y su Impacto
El uso de antibióticos en la salmonicultura chilena ha sido objeto de preocupación, con un índice de consumo de 340 gramos de antibióticos por tonelada de salmón producida en 2024, un aumento del 10.8% respecto a 2023. Esta cifra se mantiene elevada desde 2018, lo que sugiere una tendencia persistente.
La piscirickettsiosis, causada por la bacteria Piscirickettsia salmonis, es la enfermedad más prevalente y la principal causa del uso de antibióticos, representando entre el 40% y 45% de las muertes por agentes infecciosos en la industria. Eventos ambientales a finales de 2023 e inicios de 2024 propiciaron brotes de esta enfermedad, especialmente en las regiones de Los Lagos y Aysén.
El salmón del Atlántico (Salmo salar) es la especie con mayor uso de antimicrobianos, seguido por el salmón coho y la trucha arcoíris. Los principios activos más utilizados son florfenicol y oxitetraciclina. La administración es mayoritariamente oral, pero se estima que una parte significativa de estos antibióticos no es consumida por los peces y termina en el fondo marino, afectando el ecosistema y promoviendo la resistencia bacteriana.
#UsachEnMedios | Disminuyendo los antibióticos en la Acuicultura: Dra. Mónica Imarai
A pesar de la existencia de vacunas comerciales, su efectividad es debatida. El programa PROA-Salmón busca optimizar el uso de antimicrobianos, con 301.578 toneladas certificadas en 2024. Sin embargo, las concentraciones de antibióticos utilizadas siguen siendo altas, y existe el riesgo de proliferación de "superbacterias" y alteración de las comunidades microbianas del ecosistema marino.
Investigación sobre Piscirickettsia salmonis y su Interacción con Fagos
Investigaciones recientes han analizado el genoma de Piscirickettsia salmonis para comprender mejor su interacción con bacteriófagos. Se detectó la presencia de regiones de profagos (ADN viral integrado en el genoma bacteriano) en el 70% de los cromosomas analizados y en el 75% de los plásmidos. Además, se identificaron sistemas de defensa antifago, como CRISPR/Cas y sistemas de restricción-modificación.
Estos hallazgos sugieren oportunidades para el control de P. salmonis sin el uso de antibióticos, mediante la inducción de la activación de profagos para destruir la bacteria desde adentro. La comprensión de estos mecanismos de defensa también podría ser clave para desarrollar nuevas estrategias terapéuticas.
Renibacterium salmoninarum y la Enfermedad Bacteriana del Riñón (BKD)
Renibacterium salmoninarum es una bacteria Gram positiva, catalasa y oxidasa positiva, agente causal de la Enfermedad Bacteriana del Riñón (BKD). Esta patología causa mortalidad en salmónidos tanto en cultivo como en estado natural. El diagnóstico de BKD se complica por el largo tiempo de incubación de la bacteria en laboratorio.
La enfermedad se caracteriza por una infección sistémica, con síntomas externos que pueden incluir branquias pálidas, hemorragias y abdomen distendido en etapas avanzadas. Internamente, se observan lesiones macroscópicas, nodulaciones blanquecinas en riñón, hígado, corazón y bazo, esplenomegalia y ascitis.
La proteína p57 de R. salmoninarum es un importante factor de virulencia, con propiedades inmunosupresoras. La bacteria tiene la capacidad de infectar y multiplicarse dentro de células fagocíticas, evadiendo la respuesta inmune del hospedero.
Etapas de la Infección por R. salmoninarum
Se han establecido cuatro etapas de la enfermedad basadas en la detección del patógeno:
- Doble negativo: Pez sano o con infección subclínica.
- Expansión o multiplicación: Bacterias proliferando, con alta síntesis de ADN y bajos niveles de proteínas extracelulares.
- Infección o cuadro clínico: Acumulación de proteínas extracelulares y positividad para antígenos bacterianos.
- Estadio tardío: Daño renal irreversible.
La transmisión horizontal es un medio de difusión de BKD, pero la principal fuente de transmisión es vertical (padres a progenie a través de los huevos). A pesar de los programas de screening, BKD sigue siendo responsable de pérdidas significativas en el cultivo de salmónidos.
Diagnóstico y Control de BKD en Chile
En Chile, BKD está presente desde la década de los 80, caracterizándose por alta morbilidad y baja mortalidad. Es considerado un agente ubicuitario con prevalencia variable. El control se realiza mediante tratamientos con eritromicina, aunque con efecto temporal.
Se han desarrollado diversas metodologías de diagnóstico, incluyendo Nested PCR, qPCR, ELISA, IFAT, histopatología e inmunohistoquímica. Estudios serológicos han identificado dos serogrupos, y en Chile se ha detectado un solo grupo serológico mayoritario. Los análisis filogenéticos mediante secuenciación del gen 16S rRNA de aislados nacionales muestran un 100% de similitud con la cepa de referencia.
Los experimentos de vacunación han tenido progreso limitado, y la falta de protección a veces se explica por la presencia de la proteína p57. El impacto productivo de BKD en Chile no está totalmente cuantificado, pero las pérdidas por mortalidad en agua dulce y de mar, combinadas con otras enfermedades como SRS, generan costos significativos.
Desde 2005 hasta 2014, los diagnósticos positivos a R. salmoninarum aumentaron considerablemente. El salmón coho es la especie más susceptible en Chile, seguido por el salmón del Atlántico. La XI región presenta el mayor número de diagnósticos.
El control y la prevención de BKD a través del screening y durante el alevinaje son cruciales para reducir la prevalencia y generar smolts de calidad. La investigación continua busca comprender la dinámica entre P. salmonis y sus bacteriófagos para desarrollar estrategias de control innovadoras.
tags: #bacteria #en #salmon #rio #llanquihue