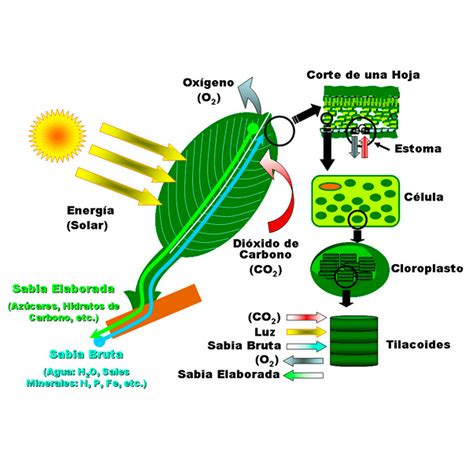
La fotosíntesis en las plantas es uno de los procesos fundamentales que mantiene el equilibrio de los ecosistemas y la vida en la Tierra. Este mecanismo bioquímico esencial implica la fabricación de nutrientes orgánicos que almacenan la energía lumínica del Sol en distintas moléculas útiles, como los carbohidratos.
Para que se produzca la fotosíntesis, es necesaria la presencia de clorofila, un pigmento sensible a la luz solar que confiere a las plantas su característico color verde. No solo las plantas acuáticas y terrestres pueden realizar la fotosíntesis, sino también otras especies como los helechos, las algas y algunas bacterias.
Fundamentos de la Fotosíntesis
Existen dos tipos principales de fotosíntesis:
- Fotosíntesis oxigénica: Es aquella que produce azúcares útiles para la planta y, a su vez, consume dióxido de carbono (CO2) y subproduce oxígeno (O2). Este es el tipo de fotosíntesis realizado por las plantas, incluyendo el poroto.
- Fotosíntesis anoxigénica: Es aquella que no produce oxígeno (O2), pero aprovecha la luz solar para romper moléculas de sulfuro de hidrógeno (H2S), común en ciertas bacterias.
El intercambio de gases que se produce durante la fotosíntesis es fundamental para los ecosistemas y para la vida. Gracias a la luz, las plantas capturan dióxido de carbono (CO2) y expulsan oxígeno durante el día. Por las noches sucede lo contrario, capturan oxígeno y liberan dióxido de carbono. Este proceso mantiene el equilibrio de los gases atmosféricos, permitiendo que el oxígeno consumido en la respiración y la combustión pueda reemplazarse.
Además de ser la base de la alimentación de muchos organismos vivos, las plantas también ofrecen propiedades medicinales, sirven como materiales de construcción (como la madera o el bambú) y protegen el suelo. La flora mantiene el equilibrio de los gases atmosféricos y es la base de alimentación de toda la pirámide de la vida. Las plantas llevan millones de años realizando este proceso que ha permitido que nuestra atmósfera se cargara de tales niveles de oxígeno que dieron lugar a la capa de ozono. Por ello, la deforestación es un impacto que amenaza la prosperidad del planeta.
Adaptaciones del Fotosistema II (PSII) frente al Estrés en Porotos
Las plantas superiores han desarrollado múltiples mecanismos de fotoprotección para utilizar eficientemente la energía absorbida y proteger su aparato fotosintético contra el daño oxidativo. Esto es particularmente crucial bajo condiciones ambientales restrictivas para el uso fotoquímico de la energía absorbida, como alta intensidad lumínica, estrés hídrico y altas temperaturas.
La mayoría de las condiciones ambientales estresantes limitan la habilidad de las plantas para utilizar la energía luminosa absorbida, produciéndose una sobreexcitación de los fotosistemas, incluso, a intensidades moderadas de luz. Consecuentemente, el daño fotooxidativo, especialmente al Fotosistema II (PSII), que es una de las estructuras más sensibles al estrés abiótico, pareciera ser una consecuencia de la actividad fotosintética y el mayor factor causante de reducciones sostenidas en la eficiencia de este proceso.
Sin embargo, los organismos fotosintéticos han desarrollado numerosos mecanismos de protección para evitar o disminuir los efectos de la absorción excesiva de luz. Entre estos, se cree que la disipación como calor del exceso de energía absorbida es el más relevante en la prevención del daño fotooxidativo del aparato fotosintético. Otros mecanismos incluyen la disminución de la intercepción de luz mediante el movimiento de hojas y cloroplastos, y la destoxificación de especies reactivas que originan daño (como clorofilas en estado de triplete excitado u oxígeno singlete).
Heterogeneidad del PSII: PSIIα y PSIIβ
La naturaleza de los mecanismos capaces de inducir la des-excitación no fotoquímica de la energía absorbida es diversa y puede facilitarse por la acidificación del lumen de los tilacoides y mediante la des-epoxidación de carotenoides xantófilos. Además, a partir de cambios conformacionales en el aparato fotosintético, componentes de los complejos PSII pueden asociarse con los PSI, traspasando energía a estos últimos, que son capaces de des-excitar pigmentos espontáneamente. Esto denota la existencia de PSII funcionales con distintas conformaciones moleculares, es decir, heterogéneos.
La heterogeneidad de los PSII está modulada por la intensidad lumínica, por la capacidad del sistema de utilizarla fotoquímicamente y por la temperatura. Una alteración observada por aumentos de temperatura es la separación de los centros de reacción (CR) de los PSII de sus complejos de antena periférica (LHCII), migrando los primeros a las regiones expuestas al estroma de los tilacoides y ubicándose en las cercanías de los fotosistemas I (PSI).
- El PSIIα es el PSII que se encuentra en su estado y ubicación normal.
- El PSIIβ es el PSII separado de su antena y ubicado en las regiones del tilacoide expuestas al estroma, cerca del PSI.
Las disminuciones en la proporción PSIIα/PSIIβ ocurren al aumentar la temperatura y se cree que corresponden a un mecanismo de protección del PSIIα contra la fotoinhibición y/o contra desbalances en la relación ATP/NADPH.
Estados de Transición: Optimización de la Distribución de Energía
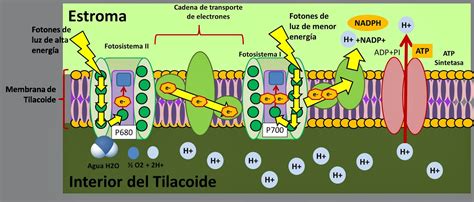
Otro mecanismo desarrollado por las plantas para optimizar la utilización de la energía capturada y regular la producción de ATP y NADPH es el llamado estado de transición. Este es considerado una respuesta a mediano plazo del aparato fotosintético frente a cambios en la proporción de excitación recibida por el PSII y PSI.
Si el PSII absorbe más energía que el PSI, se crea un desbalance en el flujo de electrones en la membrana tilacoidal, resultando en una reducción de los intermediarios del transporte de electrones. Se cree que la reducción del pool de plastoquinonas y del citocromo b6/f activa una quinasa unida a la membrana tilacoidal que fosforila las proteínas del LHCII. Esto causa la disociación de la antena (LHCII) del centro de reacción del PSII. La influencia de cargas negativas introducidas por la fosforilación permite que las membranas apiladas se desempaquen parcialmente, permitiendo la difusión del LHCII fosforilado desde la zona entre granas a la zona expuesta al estroma, cerca del PSI. Con esto se reduce el tamaño de la antena funcional del PSII, aumentando así la distribución de excitación entre los fotosistemas y disminuyendo la probabilidad de fotoinhibición.
Cuando la tasa de absorción de luz del PSI excede la del PSII, la plastoquinona y el citocromo b6/f se oxidan, induciéndose la defosforilación del LHCII por la activación de la fosfatasa unida a las membranas tilacoidales. Este fenómeno se mide como un apagamiento no fotoquímico de la fluorescencia y ocurre en condiciones de alta intensidad luminosa. La importancia de estos mecanismos, y su dependencia de los cambios de temperatura, se ha atribuido a que con frecuencia en condiciones de campo, altas temperaturas ocurren simultáneamente con elevadas intensidades luminosas y, por lo mismo, prevendrían problemas de fotoinhibición.
Investigación sobre Estrés Ambiental en Cultivares de Poroto
Los climas caracterizados por altas temperaturas y alta radiación solar normalmente se acompañan, además, por restricciones de agua. Es sabido que el déficit hídrico también afecta el proceso fotosintético, al aumentar considerablemente la resistencia estomática. Incluso, en condiciones muy severas, puede causar disfunciones en el centro de reacción de los fotosistemas y en las reacciones sucesivas del transporte fotosintético de electrones, alterar la fotofosforilación y el funcionamiento de diversas enzimas que participan en la fijación de carbono.
Un estudio se propuso analizar el efecto combinado de la temperatura y la disponibilidad hídrica sobre la proporción de los PSIIβ y los estados de transición en cuatro cultivares de poroto: Arroz Tuscola (AT), Orfeo INIA (OI), Bayos Titán (BT) y Hallado Dorado (HD).
Efectos de la Temperatura y el Estrés Hídrico
- En plantas desarrolladas en cámaras de crecimiento: La proporción de centros PSIIβ aumentó hasta en un 100% a medida que se incrementó la temperatura. Dicho efecto fue magnificado por el estrés hídrico.
- En condiciones de campo: El estrés lumínico, impuesto por la fijación de folíolos a la posición horizontal, aumentó aún más el efecto del estrés hídrico sobre la proporción de centros PSIIβ, desde un 27% en plantas regadas y hoja en posición normal, hasta un 63% en plantas estresadas y hojas forzadas a la horizontalidad. Las plantas de poroto tienden a modificar el ángulo de sus hojas para evitar la exposición directa al sol; la mantención de las hojas en una posición forzada a la horizontalidad significó que estas se expusieran a niveles de 2.200 µmol m-2 s-1 durante medio día en verano en Santiago, más del doble de la intensidad de saturación.
- En cuanto a los estados de transición: Se observó, en plantas desarrolladas a 20 ºC en cámaras de crecimiento, un aumento de estos al someter sus hojas a 15 ºC. Asimismo, temperaturas de 25 a 35 ºC indujeron aumentos en los estados de transición. El estrés hídrico, en los cultivares AT y OI, aminoró la magnitud del efecto de la temperatura, al contrario de lo observado en BT. En el cultivar HD, no se distinguió un efecto claro del estrés hídrico sobre la formación de los estados de transición inducidos por cambios en la temperatura.
Aun cuando existen diferencias en la magnitud de los cambios observados en las diferentes variedades de poroto, en todos ellos se aprecia la tendencia a modificar la estructura de los centros PSII, de manera de favorecer una menor sobreexcitación de los centros de reacción de dichos complejos, en las situaciones de estrés estudiadas.

Metodología de Estudio en Porotos
Para llevar a cabo el estudio se utilizaron los siguientes materiales y métodos:
Material vegetal
- Plantas cultivadas en cámara de crecimiento: Semillas de los cultivares Arroz Tuscola (AT), Orfeo INIA (OI), Bayos Titán (BT) y Hallados Dorados (HD) fueron sembradas en macetas plásticas. Las plantas crecieron en una cámara de ambiente controlado con temperaturas de 22/18 ºC (día/noche), un fotoperíodo de 12 h y una intensidad lumínica moderada de 500 µmol PAR m-2 s-1. La diferenciación del tratamiento hídrico se realizó regando plantas sometidas a estrés con 50 mL de agua destilada dos veces por semana (potencial hídrico foliar cercano a -1,2 MPa), mientras que las plantas sin déficit hídrico se regaron con 300 mL (potencial hídrico mayor a -0,5 MPa).
- Plantas en condiciones de campo: Se realizó un ensayo con los cultivares Orfeo INIA y Arroz Tuscola, con un diseño de bloques al azar. Las parcelas de riego normal se regaron restituyendo el 100% de la evapotranspiración (ET) diaria, mientras que las de riego restringido recibieron solo el 40% de la ET.
Mediciones
Se realizó la medición de la cinética rápida de emisión de fluorescencia. La primera hoja trifoliada de plantas cultivadas en maceta fue colocada en oscuridad por 30 min. Posteriormente, se tomaron discos de hoja de 1 cm de diámetro, a los que se les realizó una suave abrasión y fueron sumergidos en una solución de 50 µM de 3-(3,4-Dichlorophenyl)-1,1-Dimethylurea (DCMU) por 10 min, para luego proceder a la medición de la fluorescencia.
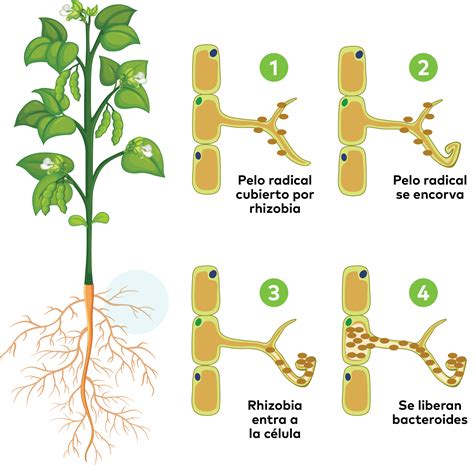
La Simbiosis Fijadora de Nitrógeno en el Poroto
La simbiosis fijadora de nitrógeno es un proceso fundamental que tiene lugar en las plantas leguminosas, como el poroto, cuando se enfrentan a condiciones de baja disponibilidad de este compuesto en el suelo. Un equipo de expertos del CONICET estudió la relación del cultivo con bacterias del suelo que favorecen la fijación de nitrógeno.
El trabajo, en su conjunto, se dio en el marco del consorcio europeo INCREASE (Colecciones Inteligentes de Recursos Genéticos de Leguminosas Alimenticias para los Sistemas Agroalimentarios Europeos), con apoyo económico de la Comisión Europea. En este contexto, el equipo liderado por Mario Aguilar, investigador del CONICET en el IBBM, evaluó la interacción entre los rizobios (bacterias fijadoras de nitrógeno) y distintos pooles genéticos de poroto.
El objetivo fue indagar sobre las características determinantes de la capacidad de fijar nitrógeno y el perfil de genes requeridos para facilitar una mejor productividad. Los pooles analizados provinieron de los distintos centros de domesticación y diseminación de este cultivo: la región mesoamericana en el que se originó; los Andes del Sur; y la región de montaña entre Ecuador y Perú. Mediante distintos análisis comparativos, los expertos pudieron determinar cuáles son los tipos de rizobios con los que las plantas de poroto común de diversas regiones de América encuentran mayor afinidad mutua.
Importancia Económica y Sostenibilidad del Cultivo de Poroto
Pese a ser una fuente importante de proteínas, hidratos de carbono, minerales y fibra, el poroto no siempre figura entre los favoritos en la dieta. Sin embargo, en Argentina, y en particular en la zona del Noroeste argentino, es un cultivo significativamente importante en materia económica. Año a año, las especies de esta leguminosa que se cultivan allí -fundamentalmente alubia y negro- tienen como destino Brasil, Argelia, Francia y España.
Estos trabajos de investigación son relevantes porque la población mundial aumentará en los próximos años, y consecuentemente se espera el aumento de la demanda de alimentación. Se revela, entonces, como meta esencial el mejoramiento de la productividad de los sectores agronómicos, que comprenda al mejoramiento de la calidad alimenticia, el desarrollo de variedades resistentes y tolerantes a estrés biótico y abiótico (es decir, de los organismos y del entorno), en el contexto de una agricultura sostenible y amigable con el medio ambiente.
tags: #proceso #fotosintesis #porotos