La beta-oxidación es un proceso metabólico crucial para la degradación de los ácidos grasos en las células del cuerpo humano. Esta vía metabólica en la cual los ácidos grasos de cadena larga se descomponen en moléculas más pequeñas para ser utilizadas como fuente de energía. Su objetivo principal es generar energía mediante la producción de acetil-CoA, que luego ingresa en el ciclo de Krebs.
Este proceso ocurre principalmente en las mitocondrias de células, como las del músculo esquelético y el corazón, y también en los peroxisomas, dependiendo del tipo de ácido graso involucrado. La beta-oxidación es un proceso catabólico en el cual los ácidos grasos sufren la eliminación sucesiva de un par de átomos de carbono en cada ciclo mediante oxidación, hasta que el ácido graso se descompone por completo en forma de moléculas de acetil-Coenzima A (acetil-CoA).
Preparación y Transporte de Ácidos Grasos a la Mitocondria
Una vez terminada la lipólisis, los ácidos grasos generados ingresan en el torrente sanguíneo para ser utilizados por el organismo en el proceso de beta-oxidación. Debido a su carácter hidrofóbico, los ácidos grasos no pueden moverse con libertad por el torrente sanguíneo y requieren un transporte adecuado.
Activación de los Ácidos Grasos
Una vez en el citoplasma, los ácidos grasos deben ser activados para poder entrar a las mitocondrias. Este proceso se llama activación de los ácidos grasos y consiste en la unión de un ácido graso a una molécula de coenzima A (CoA), formando un acil-CoA. Esta activación ocurre en la membrana externa de la mitocondria, mediante la acción catalítica de la enzima acil-CoA sintetasa (E). Los ácidos grasos (R-COO-) se activan uniéndose a una molécula de CoA-SH, con gasto de ATP, resultando acil-CoA, AMP y pirofosfato (PPi).
Transporte por la Carnitina
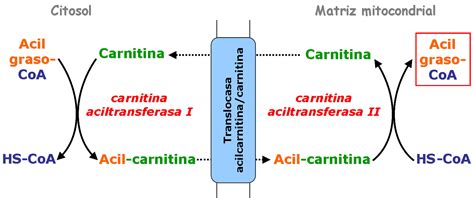
Los acil-CoA, especialmente los de cadena larga (entre 10 y 20 átomos de carbono, mayoritarios en la dieta), no pueden pasar directamente a través de la membrana mitocondrial interna. Por lo tanto, posteriormente debe usarse un transportador, la carnitina, para translocar las moléculas de acil-CoA al interior de la matriz mitocondrial. La carnitina es un derivado aminoacídico que se produce naturalmente en el hígado a partir de los aminoácidos L-metionina y L-lisina y participa en el circuito vascular reduciendo los niveles de triglicéridos y colesterol en sangre.
La enzima carnitina palmitoiltransferasa I (CPTI), situada en la membrana mitocondrial externa, elimina la coenzima A de la molécula de acil-CoA y, a la vez, transfiere y une el ácido graso a la carnitina situada en el espacio intermembrana (EIM), originando acilcarnitina. El CoA queda libre en el citosol para poder activar otro ácido graso. Los ácidos grasos de cadena más larga de 20 carbonos se oxidan en los peroxisomas, mientras que los menores de 10 carbonos entran directamente por difusión en la mitocondria.
El Proceso de Beta-Oxidación
La beta-oxidación consiste en la degradación de los ácidos grasos mediante un proceso por etapas, en cada una de las cuales se forma una molécula de acetil-CoA y dos coenzimas reducidas: FADH2 y NADH + H+. Las moléculas de acetil-CoA pueden entrar posteriormente en el ciclo de Krebs para la producción adicional de energía en forma de ATP. Para un ácido graso de cadena larga, este proceso puede repetirse muchas veces, generando grandes cantidades de ATP.
Cada acil-CoA se degrada mediante una secuencia repetitiva de cuatro reacciones enzimáticas, de forma que cada vez que el ácido se acorta en dos carbonos se va generando acetil-CoA, NADH y FADH2. Este proceso supone una visión de un ciclo en espiral ya que repite los mismos pasos, pero con diferentes sustancias procedentes del ciclo anterior.
1. Oxidación por FAD
El primer paso es la oxidación por deshidrogenación del ácido graso activado (acil-CoA graso) por FAD. La enzima acil-CoA deshidrogenasa, una flavoproteína que tiene a la coenzima Flavina-Adenina (FAD) unida covalentemente, cataliza la formación de un doble enlace entre el carbono 2 (alfa) y el carbono 3 (beta) del acil-CoA.
2. Hidratación
El siguiente paso es la hidratación (adición de una molécula de agua) del doble enlace trans entre C-2 y C-3. Esto genera un grupo hidroxilo en el tercer carbono (carbono beta).
3. Oxidación por NAD
El tercer paso es la oxidación por deshidrogenación del L-3-hidroxiacil CoA por Nicotinamida-Adenina (NAD), catalizada por la L-3-hidroxiacil CoA deshidrogenasa. Esto convierte el grupo hidroxilo del carbono beta en un grupo cetónico, perdiendo el hidrógeno del hidroxilo que es ganado por el NAD.
4. Tiólisis
El paso final es la ruptura del 3-cetoacil CoA entre C-2 y C-3 por el grupo tiol de otra molécula de CoA. Esto libera una molécula de acetil-CoA y un acil-CoA con dos carbonos menos, que vuelve a entrar en el ciclo de beta-oxidación.
🔥BETA OXIDACION de los ACIDOS GRASOS!! (FACIL) - Catabolismo de lipidos Ejercicios #bioquimica
Beta-Oxidación del Ácido Palmítico
El ácido palmítico es un ácido graso saturado que tiene 16 carbonos. Para la oxidación completa del palmitato, son necesarios 7 ciclos de beta-oxidación. Cada ciclo de beta-oxidación libera una molécula de acetil-CoA, una de FADH2 y una de NADH. Por lo tanto, a partir del ácido palmítico se obtienen los siguientes productos:
- 8 moléculas de acetil-CoA (el último ciclo libera dos unidades de acetil-CoA)
- 7 moléculas de NADH
- 7 moléculas de FADH2
Estos productos son fundamentales para la generación de energía. La baja velocidad de la oxidación del palmitato podría explicar su falta de efecto sobre la glicolisis. Además, la glucosa determina el destino metabólico del palmitato, que puede ser dirigido hacia la síntesis de lípidos a elevadas concentraciones de la hexosa, pudiendo generar mensajeros para la señalización celular.
Balance Energético del Ácido Palmítico
El cálculo del balance energético de la oxidación completa del ácido palmítico es crucial para entender su importancia como fuente de energía. Las moléculas de FADH2 y NADH producidas en la beta-oxidación y el ciclo de Krebs van a la cadena respiratoria para generar ATP. Las moléculas de acetil-CoA ingresan en el ciclo de Krebs, donde generan GTP y más moléculas de FADH2 (una por ciclo) y NADH (tres por ciclo).
Para calcular las moléculas de ATP generadas en la oxidación completa de un ácido graso, se utilizan las siguientes equivalencias energéticas modernas:
- 1 NADH produce 2.5 moléculas de ATP
- 1 FADH2 produce 1.5 moléculas de ATP
- 1 acetil-CoA (a través del ciclo de Krebs) produce 10 moléculas de ATP (considerando 3 NADH, 1 FADH2 y 1 GTP que equivale a 1 ATP).
Considerando los productos de la beta-oxidación del ácido palmítico (16 carbonos):
- 8 acetil-CoA: 8 * 10 ATP = 80 ATP
- 7 NADH: 7 * 2.5 ATP = 17.5 ATP
- 7 FADH2: 7 * 1.5 ATP = 10.5 ATP
Suma total generada: 80 + 17.5 + 10.5 = 108 ATP.
Hay que tener en cuenta que en la activación inicial del ácido graso a acil-CoA se consume el equivalente a 2 enlaces de alta energía (ATP se hidroliza a AMP y pirofosfato, lo que es el gasto de dos ATPs en términos de energía). Por lo tanto, el rendimiento neto es:
108 ATP (generados) - 2 ATP (consumidos en activación) = 106 moléculas de ATP.
Este rendimiento resalta la eficiencia de los ácidos grasos como reserva energética.