Desde hace décadas se ha determinado que los esteroides juegan un papel clave en la coordinación de las actividades del eje hipotálamo-hipófisis-gónada, además de su actividad en el control de procesos como la diferenciación y determinación sexual, metabolismo, respuesta inmune, osmoregulación, ritmos circadianos, señalización de feromonas, apareamiento y reproducción.
La reproducción involucra diversos procesos fisiológicos, controlados por órganos específicos como la glándula pineal, el hipotálamo, la hipófisis, el hígado y las gónadas. Durante las etapas de crecimiento, los niveles hormonales involucrados en la reproducción mantienen una producción relativamente baja. Una vez ocurre la madurez de la gónada, la actividad hormonal se ve influenciada por diferentes factores como la temperatura, el fotoperiodo, señales asociadas con las lluvias y otras aún poco estudiadas.
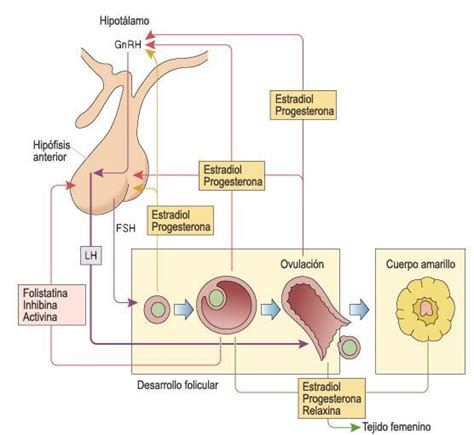
Los factores externos, como el fotoperiodo, las fases lunares y la disponibilidad de alimento, desencadenan en el cerebro del pez una serie de eventos hormonales que regulan las funciones reproductivas. En este contexto, la hormona liberadora de gonadotropinas (GnRH) actúa como un péptido hipotalámico modulador de la síntesis y liberación de gonadotropinas (GtH) como la Fsh y Lh desde la glándula pituitaria, estimulando consecuentemente la síntesis de esteroides sexuales para el crecimiento gonadal y la maduración.
Gonadotropinas Hipofisarias: FSH y LH
Los procesos de desarrollo reproductivo en un pez maduro están controlados por dos clases de gonadotropinas hipofisarias denominadas GtH I y GtH II. Dada la similitud en las secuencias entre la GtH-I (peces) y la hormona folículo estimulante (Fsh) en mamíferos, y entre la GtH-II y la hormona luteinizante (Lh), se ha adoptado el uso de los términos Fsh y Lh para referirse a GtH-I y GtH-II, respectivamente.
La Fsh está involucrada principalmente en la vitelogénesis, induciendo la captura de vitelogenina (Vtg), la cual es modulada por la síntesis folicular de estradiol (E2). Por otro lado, la principal acción de la Lh es la regulación de la maduración del oocito, la ovulación y el desove, requiriendo la síntesis previa de esteroides inductores de maduración (MIS), por lo que se la conoce como hormona madurativa.
La síntesis y secreción de ambas gonadotropinas ocurre en respuesta a la estimulación de la GnRH, la cual es suprimida por la hormona inhibidora de gonadotropinas (GnIH).
Papel de la FSH y LH en la Esteroidogénesis
La síntesis de andrógenos, principalmente 11 ketotestosterona (11-KT) y testosterona (T), se produce por la acción de la Fsh, que favorece la proliferación de espermatogonias y la espermiogénesis. En las hembras, la Lh y Fsh estimulan la producción de andrógenos a partir del colesterol en las células de la teca. Estos andrógenos, especialmente la testosterona, pasan a las células de la granulosa, donde por acción de la P450 Aromatasa, son transformados en estrógenos.
Los estrógenos, principalmente 17β-estradiol (E2), son dirigidos al torrente sanguíneo y posteriormente captados por los receptores hepáticos, estimulando la síntesis hepática de vitelogenina (Vtg).
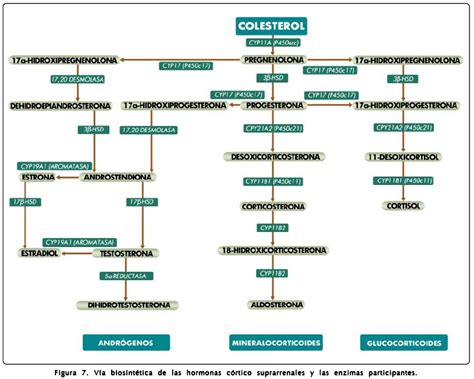
Si bien la esteroidogénesis inicia a partir del colesterol, con múltiples transformaciones mediadas por genes y enzimas caracterizadas en muchas especies de teleósteos, es importante señalar que algunas de estas enzimas y genes solo se han descrito para unos pocos teleósteos sudamericanos. Por ejemplo, la enzima hsd17b3, esencial para la síntesis de 11-Ketotestosterona, ha sido caracterizada en 46 especies de teleósteos, encontrándose en un solo Siluriforme introducido. Por su parte, la enzima hsd17b1, que convierte la estrona inactiva (E1) en estradiol (E2) activo, se ha caracterizado en 51 especies, incluyendo especies modelo como la tilapia del Nilo, anguila japonesa, pez cebra y bacalao atlántico.
Reguladores de las Gonadotropinas
Hormona Liberadora de Gonadotropinas (GnRH)
La GnRH es un péptido hipotalámico que modula la síntesis y liberación de gonadotropinas (GtH) como Fsh y Lh desde la glándula pituitaria, estimulando la síntesis de esteroides sexuales para el crecimiento gonadal y la maduración. Se ha demostrado la existencia de tres genes parálogos de GnRH: GnRH1 (catfish GnRH), GnRH2 (chicken GnRH) y GnRH3 (salmon GnRH), y al menos 14 isoformas en vertebrados. En teleósteos como Beloniformes, Perciformes y Siluriformes se han identificado GnRH1 y GnRH2, perdiendo la expresión de GnRH3. Sin embargo, existe evidencia que sugiere una compensación funcional asumida por los genes parálogos restantes.
GnRH1 y GnRH3 se expresan en el cerebro anterior ventral, el área preóptica telencefálica y el hipotálamo, siendo responsables de la regulación de la reproducción. GnRH2, en cambio, está restringida a la parte dorsal del diencéfalo y parece tener una función neuromoduladora sobre el comportamiento sexual (desove, migración), agresivo, y aparentemente actúa como factor liberador de melatonina en la glándula pineal de teleósteos.
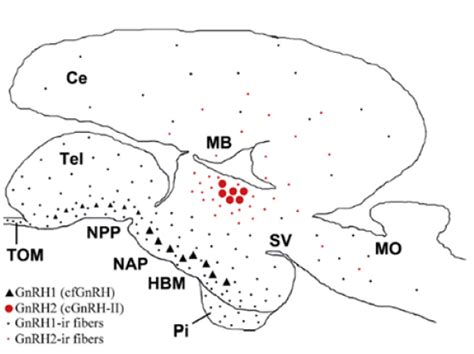
Investigaciones recientes en Silúridos sudamericanos, como Steindachneridion parahybae, han identificado la presencia y distribución de GnRH1 y GnRH2 en el cerebro. Los análisis han permitido ubicar las neuronas inmunoreactivas de GnRH1 en el hipotálamo medio basal y el área preóptica, cuyos axones inervan fuertemente la glándula pituitaria y el Pars distalis proximalis (PDP), confirmando que en los Siluriformes, la GnRH1 estimula la secreción de GtHs. En este mismo estudio, los niveles de GnRH1 se mantuvieron altos en la etapa vitelogénica, y su función moduladora en la síntesis y liberación de gonadotropinas fue confirmada, ya que la vitelogénesis está controlada por la Fsh, y la GnRH es sintetizada y liberada bajo cantidades suficientes de esteroides que modulan su actividad. Los niveles de GnRH2 variaron poco a lo largo del ciclo reproductivo, aunque se encontraron elevados en la etapa de regresión, sugiriendo un papel autocrino y/o paracrino en la maduración ovárica caracterizada por oocitos atrésicos.
Otros Neurotransmisores y Neuropéptidos Moduladores
Estudios en trucha arcoíris (Oncorhynchus mykiss) han demostrado el papel inhibitorio de la dopamina en la liberación de GnRH1. Bajo condiciones de cultivo en alta densidad, se observó inhibición sobre la Fsh hipofisaria, con una disminución significativa en los niveles de expresión de ARNm de Fsh, seguida de aumentos en los niveles de dopamina sérica.
Otros neurotransmisores como la noradrenalina, serotonina, neuropeptido Y, y las kisspectinas y sus receptores GPR54, han demostrado un papel estimulador en la liberación de GnRH. Las kisspectinas (Kiss 1 y Kiss 2) son poderosos activadores en la liberación de gonadotropinas, ya que despolarizan las neuronas de GnRH1 y estimulan la liberación de Lh y Fsh desde la pituitaria al inicio de la pubertad.
Hormona Inhibidora de Gonadotropinas (GnIH)
La GnIH es un dodecapéptido identificado inicialmente en la codorniz japonesa (Coturnix japonica) que actúa sobre las neuronas de la glándula pituitaria y la GnRH, disminuyendo la liberación y síntesis de gonadotropinas en mamíferos, aves, reptiles y anfibios.
En teleósteos, la investigación sobre la GnIH aún es objeto de estudio y debate, pero existe acuerdo sobre su papel en la regulación del eje hipotálamo-pituitaria-gónada (HPG). En hembras de Carassius auratus inyectadas con péptidos sintéticos de GnIH, se encontró una disminución significativa de los niveles de ARNm de gnrh3 y la subunidad β de Fsh (fshb) en la glándula pituitaria. GnIH también disminuyó significativamente los niveles de ARNm de la subunidad β de Lh (lhb). Cuando GnIH3 se aplicó a células primarias de cultivo de pituitaria, se observó inhibición en la síntesis de lhb y fshb estimulada por GnRH.
En carpa común (Ciprinus carpio), especie importante en acuicultura, también se ha estudiado el rol negativo del sistema GnIH/GnIHR sobre la expresión de gonadotropinas. Hembras maduras inyectadas con péptido GnIH3 presentaron cambios en los niveles de expresión de ARNm de gonadotropinas, y efectos similares se encontraron in vitro. Adicionalmente, se detectaron bajos niveles de expresión de ARNm de gnih en los meses previos al desove y un aumento significativo después del desove.
No obstante, se han reportado resultados diferentes en el salmón rojo (Oncorhynchus nerka) y tilapia (Oreochromis niloticus). En salmón rojo, se encontró un efecto en la liberación de Fsh, Lh y la hormona de crecimiento (GH) dependiente de la dosis de péptidos de GnIH. En tilapia, se encontraron efectos positivos en la liberación de Fsh y Lh in vivo e in vitro tras la inyección de GnIH2.
Investigación en la Purificación y Desarrollo de Inmunoensayos de LH
Un estudio detallado se centró en la purificación y caracterización de la hormona luteinizante (LH) de la lubina europea (Dicentrarchus labrax) y el desarrollo de un inmunoensayo específico (ELISA). La LH, secretada por la glándula hipofisaria, regula los procesos de maduración gonadal, ovulación/espermiación y puesta en vertebrados.
El proceso de purificación incluyó cromatografía en columna (gel filtración, intercambio iónico y FPLC). Las subunidades α y β de la LH se aislaron mediante rpHPLC. El peso molecular de la LH se estimó en 31 kD, y el de sus subunidades α y β en 12 y 22 kD, respectivamente. Se obtuvieron anticuerpos específicos contra la subunidad LHβ, que fueron utilizados para desarrollar un inmunoensayo tipo ELISA con una sensibilidad de aproximadamente 0.65 ng mL-1.
La validación del ensayo mostró paralelismo entre la curva patrón y muestras de plasma e hipófisis de lubina, así como con extractos hipofisarios de otras especies de peces perciformes. Mediante ELISA, se analizaron los niveles plasmáticos de LH en lubinas sometidas a tratamiento de inducción hormonal a la puesta, observándose un marcado incremento de la LH plasmática tras la inyección de diferentes dosis de GnRHa. El ELISA desarrollado se considera una herramienta sensible y precisa para cuantificar LH en muestras biológicas de lubina, fundamental para estudios sobre la endocrinología de la reproducción de esta especie.
Desnaturalización, purificación y caracterización de las proteínas
Implicaciones en Acuicultura y Investigación Futura
La investigación sobre la regulación neuroendocrina de la reproducción en peces, particularmente en el eje Hipotálamo - Pituitaria - Gónada (HPG), es crucial para el avance de la acuicultura. Se ha observado que el papel regulador de las hormonas como la Fsh y Lh puede variar significativamente según las estrategias reproductivas de la especie y las condiciones en las que se mantienen los individuos durante los estudios.
Históricamente, se ha descrito que la Fsh actúa en las fases tempranas de la reproducción, estimulando la biosíntesis de estradiol y testosterona, mientras que la Lh está implicada en las fases finales. Sin embargo, en especies con desarrollo asincrónico y desove múltiple, la Lh se produce durante todo el ciclo en paralelo con la Fsh, sugiriendo la necesidad de ambas hormonas en la gametogénesis y maduración final.
Investigaciones recientes en especies con comportamiento migratorio reproductivo han evidenciado una disminución significativa en la expresión del ARNm de la subunidad β de Lh (lhb) cuando los individuos son mantenidos en cautiverio. Esto podría ocasionar disfunciones reproductivas en los sistemas de cultivo actuales, representando un desafío importante para la acuicultura contemporánea.