El resultado agronómico del cultivo de poroto deriva de la compleja interacción entre un conjunto de factores. Todos ellos en mayor o menor medida son susceptibles de ser manejados con el propósito de generar las condiciones de crecimiento y desarrollo que permitan optimizar el nivel de producción unitaria. El producto de las eficiencias individuales de una serie de prácticas agronómicas permite estimar el rendimiento del conjunto.
Importancia y Características del Cultivo de Poroto
El poroto es un cultivo de gran relevancia, con Argentina destinando casi la totalidad de su producción (90%) a la exportación, lo que genera un gran interés del productor porotero en las tecnologías disponibles para su cuidado. Se trata de un cultivo propio de Salta, Jujuy y Tucumán, ganando terreno en el noroeste argentino, región que le ofrece condiciones óptimas para su desarrollo gracias a su clima seco y gran amplitud térmica.
En esta región se cultivan especies como Phaseolus L. y Vigna Savi, de las que provienen variedades como porotos adzuki, alubia, blanco, negro, colorado, manteca, mung, pallar y poroto tape o caupí. El poroto se caracteriza por ser un cultivo de ciclo muy corto, de 90 a 120 días. Sus plantas poseen un sistema radical superficial, donde el 95% de las raíces activas se encuentran en los primeros 20 cm de profundidad del suelo. Esta particularidad lo hace muy exigente en condiciones de alta disponibilidad de nutrimentos en la solución edáfica. Es sensible a la baja disponibilidad de potasio, calcio, zinc, manganeso, hierro y molibdeno, así como a condiciones adversas de salinidad y toxicidad específica de boro, cloro y sodio.
Aunque exigente en nitrógeno, su limitación se mitiga por su capacidad de fijarlo del aire mediante simbiosis con bacterias del género Rhizobium. Esta fijación simbiótica ocurre espontáneamente en suelos con rizobios autóctonos, aunque a menudo con baja eficiencia. En cultivos conservacionistas de suelo, la eficiencia de fijación de nitrógeno es mayor, incrementando las probabilidades de éxito de la inoculación. En el caso del frejol (Phaseolus vulgaris L.), la temperatura mínima para que ocurra su germinación es de 8°C.

La Temperatura como Factor Determinante en el Manejo Agronómico
La siembra y la cosecha son operaciones fundamentales que el agricultor realiza anualmente. Para optimizar el rendimiento, es crucial analizar las particularidades del clima, el sistema de producción y los factores que limitan el cultivo de poroto en cada zona.
Importancia de la Fecha de Siembra
La fecha de siembra es esencial, ya que permite optimizar la coincidencia de las condiciones climáticas de cada zona con las necesidades fisioecológicas de las distintas etapas de desarrollo a lo largo del ciclo del cultivo, maximizando así la expresión del potencial genético.
Manejo Preventivo y la Temperatura
El conocimiento de las temperaturas, heladas, humedad relativa y heliofanía de la zona es una herramienta imprescindible para el manejo preventivo de enfermedades. Este manejo consiste en identificar la etapa y fase del cultivo en la que puede aparecer la enfermedad, el nivel de tolerancia o susceptibilidad de la variedad utilizada y las condiciones ambientales que el patógeno necesita para desarrollarse.
Mecanismos Fisiológicos de Adaptación al Estrés Térmico y Lumínico
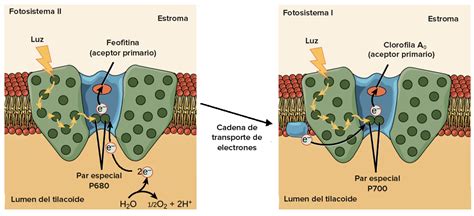
La mayoría de las condiciones ambientales estresantes limitan la capacidad de las plantas para utilizar la energía luminosa absorbida, lo que puede llevar a una sobreexcitación de los fotosistemas, incluso con intensidades de luz moderadas. El daño fotooxidativo, particularmente al Fotosistema II (PSII), una de las estructuras más sensibles al estrés abiótico, es una consecuencia de la actividad fotosintética y un factor importante en la reducción de su eficiencia.
Las plantas han desarrollado numerosos mecanismos de protección para mitigar los efectos de la absorción excesiva de luz, incluyendo: disminuir la intercepción de luz mediante el movimiento de hojas y cloroplastos, disipar como calor la energía absorbida en exceso, y/o destoxificar especies reactivas que causan daño. Se cree que la disipación de calor es el mecanismo más relevante en la prevención del daño fotooxidativo del aparato fotosintético.
La Heterogeneidad del Fotosistema II (PSII)
La heterogeneidad de los PSII está modulada por la intensidad lumínica, la capacidad del sistema para utilizarla fotoquímicamente y la temperatura. Una alteración observada con aumentos de temperatura es la separación de los centros de reacción (CR) de los PSII de sus complejos de antena periférica (LHCII), migrando los primeros a las regiones expuestas al estroma de los tilacoides y ubicándose cerca de los fotosistemas I (PSI).
- El PSII en su estado y ubicación normal se denomina PSIIα.
- El PSII separado de su antena y ubicado en las regiones del tilacoide expuestas al estroma se denomina PSIIβ.
Los centros β han sido identificados mediante el análisis de la cinética rápida de emisión de fluorescencia. Las disminuciones en la proporción PSIIα/PSIIβ que ocurren al aumentar la temperatura se interpretan como un mecanismo de protección del PSIIα contra la fotoinhibición y/o contra desbalances en la relación ATP/NADPH.
Los Estados de Transición
Otro mecanismo que las plantas han desarrollado para optimizar el uso de la energía capturada y regular la producción de ATP y NADPH es el llamado estado de transición. Esta es una respuesta a mediano plazo del aparato fotosintético frente a cambios en la proporción de excitación recibida por el PSII y PSI. Si el PSII absorbe más energía que el PSI, se produce un desequilibrio en el flujo de electrones en la membrana tilacoidal, resultando en una reducción de los intermediarios del transporte de electrones.
Se postula que la reducción del pool de plastoquinonas y del citocromo b6/f activa una quinasa unida a la membrana tilacoidal que fosforila las proteínas del LHCII. Esto provoca la disociación de la antena (LHCII) del centro de reacción del PSII. Las cargas negativas introducidas por la fosforilación permiten que las membranas apiladas se desempacen parcialmente, facilitando la difusión del LHCII fosforilado desde la zona entre granas a la zona expuesta al estroma, cerca del PSI. Esto reduce el tamaño de la antena funcional del PSII, aumentando la distribución de excitación entre los fotosistemas y disminuyendo la probabilidad de fotoinhibición.
Cuando la tasa de absorción de luz del PSI excede la del PSII, la plastoquinona y el citocromo b6/f se oxidan, induciendo la desfosforilación del LHCII por la activación de la fosfatasa unida a las membranas tilacoidales. Este fenómeno se mide como un apagamiento no fotoquímico de la fluorescencia y ocurre en condiciones de alta intensidad luminosa. La importancia de estos mecanismos y su dependencia de los cambios de temperatura se atribuye a que, en condiciones de campo, altas temperaturas a menudo coinciden con elevadas intensidades luminosas, lo que permite prevenir problemas de fotoinhibición.
Interacción con el Estrés Hídrico y Lumínico
Los climas con altas temperaturas y alta radiación solar frecuentemente se acompañan de restricciones de agua, ya sea por veranos secos o por una evapotranspiración que supera la capacidad de mantener potenciales hídricos estables. El déficit hídrico afecta el proceso fotosintético al aumentar la resistencia estomática y, en condiciones severas, puede causar disfunciones en el centro de reacción de los fotosistemas, alterar el transporte fotosintético de electrones y la fotofosforilación, y afectar el funcionamiento de enzimas de fijación de carbono.
Las observaciones anteriores que correlacionan aumentos de temperatura con la formación de centros tipo β, interpretadas como un mecanismo de fotoprotección, sugieren que estos ajustes también se relacionan con otros factores estresantes como el déficit hídrico y las altas intensidades lumínicas. Las plantas de poroto tienden a modificar el ángulo de sus hojas para evitar la exposición directa al sol. Mantener las hojas en una posición forzada a la horizontalidad puede significar una exposición a niveles de luz de 2.200 µmol m-2 s-1 durante medio día en verano, en comparación con los 300 a 400 µmol m-2 s-1 PAR en una posición normal, lo que representa más del doble de la intensidad de saturación.
Estudio Experimental sobre la Respuesta del Poroto a Estreses Combinados
Un estudio realizado (De Simone, M.E. y V.N.) se propuso analizar el efecto combinado de la temperatura y la disponibilidad hídrica sobre la proporción de los PSIIβ y los estados de transición en cuatro cultivares de poroto.
Metodología del Estudio
Material Vegetal y Condiciones en Cámara de Crecimiento
Se utilizaron semillas de poroto de los cultivares Arroz Tuscola (AT), Orfeo INIA (OI), Bayos Titán (BT) y Hallado Dorado (HD), procedentes de la colección de germoplasma del Laboratorio de Leguminosas de Grano de la Universidad de Chile. Las plantas se cultivaron en macetas con una mezcla de arena, tierra de hojas y suelo (1:1:1) en una cámara de ambiente controlado con temperaturas de 22/18 ºC (día/noche), un fotoperíodo de 12 horas y una intensidad lumínica moderada de 500 µmol PAR m-2 s-1, usando lámparas de sodio. Las plantas se regaron periódicamente con solución nutritiva Hoagland número 2 hasta alcanzar el estado de primera hoja trifoliada.
Para la diferenciación del tratamiento hídrico, las plantas sometidas a estrés hídrico se regaron con 50 mL de agua destilada dos veces por semana, hasta alcanzar un potencial hídrico foliar cercano a -1,2 MPa. Las plantas sin déficit hídrico, en cambio, se continuaron regando con 300 mL de agua destilada con la misma frecuencia, manteniendo un potencial hídrico foliar mayor a -0,5 MPa.
Condiciones de Campo
Se llevó a cabo un ensayo en condiciones de campo con los cultivares Orfeo INIA y Arroz Tuscola, empleando un diseño de bloques al azar con parcela dividida. Cada bloque incluyó dos parcelas (riego normal y riego restringido), que a su vez se dividieron en subparcelas según el cultivar. El riego de las parcelas de riego normal restituyó el 100% de la evapotranspiración (ET) diaria, mientras que las de riego restringido recibieron solo el 40% de la ET, una vez que las plantas alcanzaron el estado de segunda hoja trifoliada.
Mediciones
La medición de la cinética rápida de emisión de fluorescencia se realizó en discos de la primera hoja trifoliada, previamente colocados en oscuridad y tratados con 50 µM de 3-(3,4-Dichlorophenyl)-1,1-Dimethylurea (DCMU).
Resultados y Observaciones Clave
En plantas desarrolladas en cámaras de crecimiento, la proporción de centros PSIIβ aumentó hasta en un 100% a medida que se incrementó la temperatura. Este efecto fue magnificado por el estrés hídrico en estas plantas.
En condiciones de campo, el estrés lumínico causado por la fijación de folíolos a una posición horizontal intensificó aún más el efecto del estrés hídrico sobre la proporción de estos centros. Se observó un aumento desde un 27% en plantas regadas con hojas en posición normal, hasta un 63% en plantas estresadas con hojas forzadas a la horizontalidad.
En cuanto a los estados de transición, se apreció un aumento en plantas desarrolladas a 20 ºC en cámaras de crecimiento al someter sus hojas a 15 ºC. Del mismo modo, temperaturas entre 25 y 35 ºC también indujeron incrementos en los estados de transición. El estrés hídrico aminoró la magnitud del efecto de la temperatura en los cultivares Arroz Tuscola y Orfeo INIA, mientras que en Bayos Titán el efecto se incrementó. En el cultivar Hallado Dorado, no se distinguió un efecto claro del estrés hídrico sobre la formación de los estados de transición inducidos por cambios de temperatura.
A pesar de las diferencias en la magnitud de los cambios observados entre las distintas variedades de poroto, en todos los casos se evidenció una tendencia a modificar la estructura de los centros PSII. Este ajuste fisiológico busca favorecer una menor sobreexcitación de los centros de reacción de dichos complejos en las situaciones de estrés estudiadas, actuando como un mecanismo de protección.