La membrana plasmática de la levadura es una estructura proteo-lipídica que delimita al protoplasto y protege la célula, proporcionando un entorno estable en su interior. Es una barrera altamente selectiva, permitiendo la entrada de oxígeno y nutrientes, y la excreción de productos de desecho. La composición química de las membranas les confiere propiedades esenciales para sus funciones, incluyendo la semipermeabilidad, la fluidez y la heterogeneidad lateral.
Composición y Estructura de la Membrana Plasmática
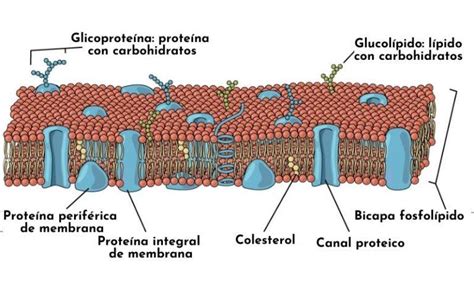
La membrana plasmática es una bicapa de fosfolípidos hidrófobos intercalados con proteínas globulares (permeasa) que permiten el intercambio de materiales. Incluye una variedad de compuestos lipídicos como:
- Fosfolípidos: que otorgan fluidez a la estructura.
- Esteroles: (incluidos los ergosteroles y el zimosterol, así como otros esteroles menores), críticos para la división celular. La levadura requiere oxígeno (proporcionado por la oxigenación inicial del mosto) para sintetizar estos compuestos. Los esteroles son derivados poliisoprenoides.
La bicapa lipídica comprende una cabeza hidrófila que permanece en la capa acuosa y una cola hidrófoba que excluye el agua. También hay ATPasa hidrolasa para generar ATP y proporcionar un gradiente de protones electroquímico necesario para el transporte de medios. A microscopio óptico, se recurre a la tinción con Azul Victoria para visualizarla. El modelo de fluido de Singer y Nicholson es ampliamente aceptado para describir su estructura, como un mosaico fluido de lípidos, igualmente dotados de una rápida movilidad.
Propiedades Fundamentales de la Membrana Plasmática
1. Semipermeabilidad
Esta propiedad es consecuencia del ambiente hidrófobo interno de la membrana, creado por las cadenas de ácidos grasos de los lípidos, que es difícil de cruzar por moléculas con carga eléctrica neta. Esta capa hidrofóbica impide la libre difusión de moléculas y permite a las membranas crear compartimentos intracelulares con contenidos químicos específicos, y mantener separados el medio intracelular del extracelular. Sin embargo, la permeabilidad es selectiva, es decir, no todas las moléculas tienen la misma dificultad para cruzar la membrana por difusión pasiva.
Factores que Influyen en la Permeabilidad
- Polaridad y tamaño de la molécula: Moléculas pequeñas sin carga (CO2, N2, O2) o con alta solubilidad en grasas (etanol) cruzan las membranas sin dificultad por difusión pasiva.
- Moléculas con carga eléctrica: La permeabilidad es menor para moléculas que tienen cargas eléctricas pero que son globalmente neutras (agua o glicerol). Existen acuaporinas en determinadas membranas que facilitan el cruce del agua.
- Moléculas grandes y cargadas: Es aún menor la capacidad para atravesar la membrana para moléculas grandes con cargas pero globalmente neutras, como la glucosa. Sin embargo, es altamente impermeable a los iones y a las moléculas que tienen carga neta.
La desigual distribución de iones y moléculas entre ambos lados de la membrana es la base para la creación de gradientes químicos y eléctricos. La medida de esa diferencia de concentración de cargas es el potencial de membrana, usado para funciones celulares como la síntesis de ATP o la transmisión del impulso nervioso. La semipermeabilidad de la membrana también permite la ósmosis, es decir, el flujo de agua hacia donde haya mayor concentración de solutos.
La permeabilidad de las membranas depende de la composición de lípidos. Membranas más fluidas suelen ser más permeables, y viceversa. Por ejemplo, el aumento de la concentración de colesterol en la membrana plasmática suele hacer que las membranas aumenten su hidrofobicidad, volviéndolas más impermeables. Esto es diferente en diferentes tipos de membranas dentro de la propia célula.
2. Fluidez
La fluidez es la capacidad de una molécula que forma parte de una membrana para desplazarse lateralmente por ella. Las membranas son fluidas, láminas de grasa donde las moléculas se encuentran en un estado de líquido viscoso.
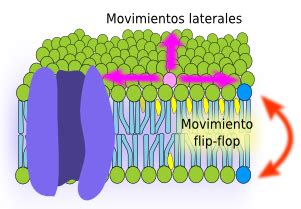
Movimientos de los Lípidos en la Membrana
Un glicerofosfolípido en la hemicapa externa de la membrana plasmática tiene dos posibilidades de movimiento:
- Movimiento lateral: Mucho más frecuente, donde se desplaza entre las moléculas contiguas. Una molécula lipídica puede recorrer 30 micras en unos 20 segundos por difusión pasiva lateral.
- Movimiento "flip-flop": Salto a la hemicapa interna. Es muy infrecuente debido a que las cabezas polares de los lípidos se encuentran con la barrera de las cadenas de ácidos grasos. Sin embargo, el colesterol posee la capacidad de hacer movimientos "flip-flop" con relativa facilidad.
Factores que Afectan la Fluidez
- Composición química: Menor longitud o mayor cantidad de enlaces insaturados de las cadenas de ácidos grasos hacen que las membranas sean más fluidas.
- Colesterol: Su efecto depende de la temperatura y composición lipídica. Inhibe el paso a estado de gel sólido (menos fluido) pero disminuye la flexibilidad de los ácidos grasos. A bajas temperaturas, el aumento de colesterol favorece la fluidez. Un efecto adicional del incremento de la concentración de colesterol es que aumenta la hidrofobicidad, es decir, las membranas se vuelven más impermeables.
- Asimetría de las membranas: La diferente composición entre las dos hemicapas puede generar diferencias en la fluidez. Se conocen dos fases: líquido ordenado (menos fluido) y líquido desordenado (más fluido). La hemicapa externa se cree más propensa a estar en fase líquido ordenado, y la interna en líquido desordenado.
Las células pueden alterar la fluidez de sus membranas modificando su composición química. Por ejemplo, en las bacterias, la adaptación de la fluidez a las condiciones ambientales se debe a cambios en la cantidad de saturación y longitud de los ácidos grasos, y a la variación en la concentración de glicerofosfolípidos como la fosfatidiletanolamina. Las mitocondrias deben crear una barrera permeable en su membrana interna para generar un gradiente de protones estable.
3. Heterogeneidad Lateral
Aunque la fluidez podría sugerir una distribución aleatoria de moléculas, la membrana es heterogénea, con dominios de diferentes composiciones moleculares debido a restricciones en la movilidad lateral. A escalas inferiores a los 200 nm, aparecen microdominios de membrana, de unos 60 a 100 nm de tamaño.
Mecanismos de Restricción del Movimiento
- Interacción con citoesqueleto o matriz extracelular: Interacción directa de moléculas de la membrana con moléculas del citoesqueleto o la matriz extracelular.
- Interacciones moleculares: Entre las propias moléculas de la membrana.
- Cambios de densidad: Menor fluidez local.
- Concentración desigual de cargas eléctricas.
- Grado de curvatura o espesor de la membrana.
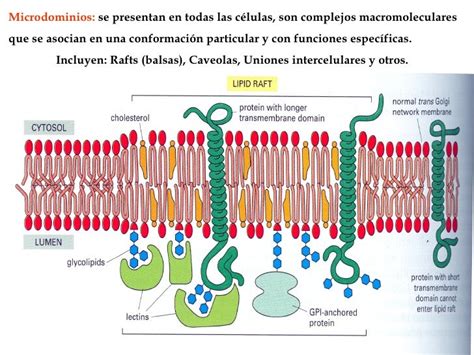
Microdominios y Balsas Lipídicas
Las interacciones entre lípidos (especialmente esfingolípidos y colesterol) pueden crear zonas de mayor densidad lipídica. Estas asociaciones, denominadas "balsas de lípidos" (lipid rafts), son muy abundantes y dinámicas, formando un mosaico de dominios más densos que viajan entre los glicerofosfolípidos, más fluidos. Las balsas son pequeñas (10-200 nm) y dinámicas, y se postula que la alta concentración de ciertos tipos de lípidos en ellas crea un ambiente químico propicio para reacciones o interacciones moleculares específicas.
En la hemicapa interna de la membrana plasmática se forman otros microdominios por interacciones electrostáticas entre proteínas con regiones citosólicas básicas y/o cationes divalentes, y las cabezas polares de los lípidos cargadas negativamente. También hay agrupaciones de lípidos fosfatidilinositol en torno a proteínas.
Restricciones en la Movilidad de Proteínas
Las proteínas integrales o asociadas a la membrana pueden interaccionar entre sí y ensamblarse en estructuras macromoleculares que favorezcan la transmisión de señales, reconocimiento celular, activación enzimática, movimiento celular, etc. Las proteínas integrales de membrana tienen numerosas restricciones a la movilidad, principalmente por interacciones de sus dominios intra y extracelulares con moléculas del citoesqueleto y de la matriz extracelular, respectivamente.
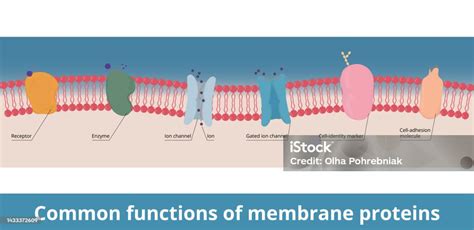
La Membrana Plasmática como Barrera Selectiva: Transporte de Sustancias
La membrana plasmática protege a la célula y transporta nutrientes hacia adentro y sustancias tóxicas hacia afuera. Es el lugar de anclaje de proteínas que le permiten interactuar con otras células, siendo estas proteínas glicoproteínas o lipoproteínas. Cada célula está unida por esta membrana que actúa como "guardián de la puerta" para mantener la homeostasis celular.
Transporte de Solutos
La membrana es una barrera que no deja atravesar solutos a partir de un determinado tamaño, dependiendo del grosor y tamaño de los poros. Las moléculas que no cruzan las membranas libremente son interesantes para las células, ya que la variación de sus concentraciones a un lado u otro puede actuar como señales o herramientas. Para ello, se han desarrollado proteínas transmembrana que permiten selectivamente el paso de estas sustancias, unas para generar el gradiente y otras para romperlo.
Sistemas de Transporte en la Membrana Plasmática
Las células de levadura, al igual que otros microorganismos, han desarrollado diversos mecanismos para captar nutrientes y eliminar desechos a través de su membrana plasmática.
1. Difusión Pasiva
Algunas moléculas pueden cruzar la membrana por difusión pasiva, sin gasto de energía, siguiendo su gradiente de concentración. Esto es aplicable a moléculas pequeñas sin carga, gases como el O2, N2, o moléculas con alta solubilidad en grasas como el etanol. La velocidad de este transporte depende del gradiente de concentración.
2. Difusión Facilitada
La difusión facilitada es un proceso pasivo en el que proteínas transportadoras o canales facilitan el paso de moléculas a través de la membrana. Aunque no requiere gasto de energía directa, se basa en la existencia de un gradiente de concentración. Un ejemplo de esto son las acuaporinas que facilitan el transporte de agua.
3. Transporte Activo
Este tipo de transporte permite el movimiento de sustancias en contra de un gradiente de concentración, lo que requiere un gasto energético. Existen dos tipos principales de transporte activo:
a) Transporte Activo Primario (Bombas)
Utiliza directamente la energía del ATP (o de otro compuesto de alta energía) para mover sustancias. Un ejemplo lo constituye la ATP-sintasa de eubacterias.
b) Transporte Activo Secundario (Cotransporte)
Utiliza la energía almacenada en un gradiente electroquímico preexistente (generado por transporte activo primario) para mover otra sustancia. Puede ser:
- Simporte: Ambas sustancias se mueven en la misma dirección.
- Antiporte: Las sustancias se mueven en direcciones opuestas.
4. Sistemas de Translocación de Grupo (PTS - Phosphotransferase Systems)
Estos sistemas, que se conocen en muchos ejemplos en eubacterias y arqueas, implican la modificación química de la sustancia transportada, normalmente por unión covalente con un grupo químico (p. ej., fosfato). Esto permite que el azúcar entre en la célula y se convierta directamente en un intermediario de su ruta metabólica. En E. coli, el sistema de fosfotransferasa es constitutivo y transfiere un grupo fosfato a la enzima-IIB específica con la que se asocia para el mismo nutriente.
🔥 ¿Qué es el ATP? - Adenosín trifosfato | Biología Desde Cero
5. Transportadores ABC (ATP-binding Cassette)
Estos transportadores son una superfamilia de proteínas presentes en todos los reinos de la vida, aunque con variantes en cada uno de ellos. Son bombas impulsadas por ATP que transportan una amplia gama de sustratos, desde iones hasta macromoléculas, a través de la membrana. Se conocen muchos ejemplos en eubacterias y arqueas. En el caso de un sistema ABC en enterobacterias (como E. coli), la hidrólisis de ATP por el dominio de unión a ATP (ATP-binding cassette) genera la sigla "ABC".
Secreción de Proteínas a través de la Membrana
Las proteínas destinadas a insertarse en las envueltas bacterianas o a ser excretadas al medio requieren sistemas de secreción especializados.
1. Sistema Sec (General Secretion Pathway)
Este sistema es el más común y se encarga del transporte de proteínas plegadas o parcialmente plegadas a través de la membrana plasmática. Las proteínas secretadas llevan una "señal" de unos 20-30 aminoácidos, que las guía hacia el complejo de proteínas de membrana SecYEG, el cual tiene un canal interior de unos 20-30 Å. Este sistema es análogo al sistema translocador del retículo endoplásmico de eucariotas y se encuentra en bacterias, arqueas y los cloroplastos, pero no en mitocondrias ni en eucariotas.
2. Sistema Tat (Twin Arginine Translocation)
Este sistema es menos común y se especializa en el transporte de proteínas ya plegadas (que pueden contener cofactores, como grupos FeS, molibdopterina, cofactores nucleotídicos, etc.) a través de la membrana. Las proteínas destinadas al sistema Tat llevan una secuencia de señal especial con un motivo "twin arginine". Hay tres tipos de proteínas de membrana Tat: TatA, TatB y TatC. El canal formado por TatA puede tener un diámetro variable de hasta 145 Å.
Componentes Celulares Adicionales en Levadura
Pared Celular
La pared celular proporciona a la célula resistencia mecánica para resistir cambios de presión osmótica y preservar la forma e integridad de la levadura durante el crecimiento y la división celular. Representa el 26-32% de la célula y está compuesta por polisacáridos (30-60%, polímeros de betaglucano y manano), proteínas (15-30%) y lípidos (5-20%), con una pequeña cantidad de quitina. La proteína ligada a los manano-oligosacáridos (MOS) se denomina complejo manoproteico.
Floculación
La floculación es una característica importante de la levadura que se produce al final de la fermentación y es un mecanismo cooperativo para proteger la célula contra tensiones (pH, bajas temperaturas, falta de nutrientes, alto contenido de etanol). Permite que las células reduzcan el impacto de un ambiente adverso al aglutinarse y asentarse por sedimentación. Es útil para los cerveceros para separar la levadura de la cerveza al final de la fermentación.
Fenotipos de Floculación
- Levadura Ale: Flocula mediante la acción de lectinas controladas por el fenotipo Flo 1, que se unen a los azúcares manosa. La floculación es inducida por etanol y puede ser inhibida por azúcares manosa, pero no depende de iones calcio. Las cepas de levadura Ale tienen macromoléculas hidrófobas en la superficie de la pared celular que se unen a burbujas de dióxido de carbono, permitiendo que los grumos de levadura floten.
- Levadura Lager: Flocula mediante la acción de lectinas controladas por el fenotipo NewFlo, que se unen a los azúcares de manosa y glucosa. Es inhibida por la presencia de manosa, glucosa, maltosa y sacarosa. La floculación ocurre solo al final de la fermentación, cuando los azúcares simples ya no están presentes, y requiere la presencia de iones de calcio solubles. Las células Lager no se unen al dióxido de carbono, sino que se depositan por gravedad.
Factores que Favorecen la Floculación
- Oxigenación: Una baja aireación del mosto puede resultar en floculación temprana o incompleta.
- Temperatura: Diferentes cepas floculan bajo diferentes condiciones de temperatura.
- pH: El rango óptimo es de 3,5 a 4,8.
- Concentración de etanol: Tanto los aumentos como las disminuciones en los niveles de etanol pueden mejorar la floculación.
- Tasa de inoculación: Tasas más altas pueden producir poblaciones con un mayor porcentaje de células más viejas que floculan antes.
- Generación: La floculencia de una cepa de levadura cambia con el relanzamiento en serie debido a cambios en la composición de la pared celular y variación genética.
- Trub: El aumento de los niveles de trub puede incrementar la floculencia en algunas cepas.
- Azúcares de mosto y sales minerales: La manosa bloquea la floculación en cepas Ale y Lager. Se requieren iones de calcio para la floculación de levadura Lager.
- Lavado ácido: Puede usarse para romper la floculación.
Espacio Periplásmico
El periplasma no es un orgánulo, sino un espacio donde se almacena el glucógeno, el primer carbohidrato utilizado al inicio de la fermentación. Es el lugar de importantes enzimas de la levadura, como los fosfatos ácidos y la invertasa (que descompone la sacarosa en glucosa y fructosa para una adsorción más fácil). En las cepas de levadura Lager, también está presente la enzima melibiosa, que puede degradar la melibiosa del azúcar.
Citoplasma
El espacio debajo de la membrana plasmática contiene material semilíquido llamado citosol, en el que están suspendidos todos los orgánulos intracelulares. Contiene sales disueltas, metabolitos celulares y enzimas, y es el lugar de muchas reacciones bioquímicas, como la vía glucolítica.
Mitocondrias
Las mitocondrias son la "potencia" de la célula. En condiciones aeróbicas y con baja concentración de azúcar, contienen las vías responsables de generar energía en forma de ATP (ciclo de Krebs y cadena de transporte de electrones). Tienen una doble membrana, siendo la interna con pliegues llamados crestas, donde ocurren algunos pasos de la respiración celular. Realizan funciones importantes durante el crecimiento anaeróbico, como la síntesis y desaturación de ácidos grasos y lípidos, biosíntesis de ergosterol, respuestas al estrés, y la producción de algunos componentes del sabor.
Tráfico Celular y Partículas de Almacenamiento
Cuando los nutrientes son abundantes, las células de levadura almacenan el exceso de lípidos (esteroles y ácidos grasos) y las reservas de carbohidratos (glucógeno) en partículas del citoplasma. Los lisosomas, presentes en el citoplasma, contienen enzimas hidrolíticas que digieren macromoléculas y degradan materiales extraños o la propia célula al morir.
Retículo Endoplásmico (RE)
Es un sistema de membranas huecas que se extiende por todo el citoplasma. Hay dos tipos:
- RE liso: Involucrado en la síntesis de lípidos, metabolismo de carbohidratos y desintoxicación.
- RE rugoso: Estructuras aplanadas con ribosomas adheridos, transporta proteínas a su ubicación dentro de las células.
Ribosomas
Los ribosomas son los sitios donde se sintetizan proteínas como las enzimas. Se encuentran en grandes cantidades en el citoplasma (adheridos al RE rugoso) y dentro de las mitocondrias. Están formados por ARN y proteínas.
Núcleo
La mayor parte del ADN o genoma de la célula (16 cromosomas en la levadura de cerveza) se almacena en el núcleo, el cual está unido por una membrana perforada por poros. Dentro del núcleo se encuentra el nucleolo, donde se sintetizan los ribosomas.
Vacuola
La vacuola participa principalmente en el reciclaje de moléculas grandes (como proteínas redundantes en aminoácidos), y sirve como cámara de almacenamiento y "papelera". Tiene una membrana lipídica para mantener el agua salada fuera del citoplasma.
Cicatriz de Yema
Cada vez que una célula de levadura se reproduce por gemación, deja una cicatriz característica y permanente en la superficie de la pared celular, rica en quitina. La nueva célula hija también tiene una "cicatriz de nacimiento" donde estaba unida a la célula madre, que es menos rica en quitina y desaparece gradualmente.
Red Citoesquelética
El citoesqueleto es una serie de microtúbulos y microfilamentos que proporcionan estructura y organización a la célula, dándole la capacidad de ordenar sus orgánulos. Son varillas huecas compuestas de tubulina, importantes en los movimientos cromosómicos, la motilidad celular, el movimiento de los orgánulos y el mantenimiento de la forma celular.
tags: #membrana #plasmatica #como #barrera #selectiva #levadura