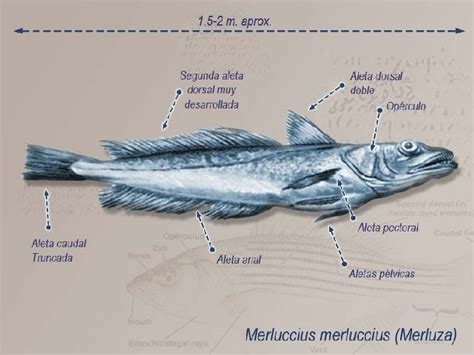
La merluza común (Merluccius gayi gayi) fue por décadas una de las especies más consumidas por chilenos y chilenas. Popularmente conocida como “la pescada”, es un pez demersal, lo que significa que vive en las profundidades del mar. Su distribución abarca las regiones de Coquimbo y Los Lagos en Chile. El tamaño promedio de esta especie es de 35 centímetros, siendo las hembras algo más grandes. Es un depredador bentónico universal, alimentándose de calamares, diversos crustáceos como los langostinos, e incluso otras merluzas.
Historia de la Pesca y Desafíos de Conservación
La historia de la pesca de la merluza es bastante amplia. Su explotación a nivel industrial comenzó en la segunda mitad del siglo XX y, desde entonces, la disponibilidad del recurso ha variado significativamente. Desde 2005, se ha observado un agotamiento paulatino del recurso, explicado por diversas razones.
En primer lugar, la institucionalidad en el país no cuenta con las herramientas necesarias para una fiscalización sistematizada y efectiva del cumplimiento de las cuotas, lo que dificulta frenar la pesca ilegal. En segundo lugar, ha habido una subestimación de la biomasa total, lo que ha llevado a pescar más de lo correspondiente para mantener una biomasa equilibrada del recurso. Por ejemplo, la cuota anual impuesta para 2020 fue de 37.900 toneladas, mientras que el pico de extracción se registró en 2001 con 122.000 toneladas. Para prevenir el colapso de la merluza común, se ha implementado una veda que se aplica durante todo el mes de septiembre.

Clasificación Taxonómica y Distribución de Merluccius gayi
Evolución y Confusión Taxonómica del Género Merluccius
El género Merluccius (Rafinesque 1810) es el más diverso de la familia Merlucciidae, contando con 12 especies, a las cuales se sumaría M. hernandezi (Mathews 1985). Este género ha sido considerado conflictivo en cuanto a sus relaciones taxonómicas y filogenéticas (Inada 1989, Paterson & Rosen 1989). Un ejemplo es la especie M. gayi (Guichenot 1848), que inicialmente fue identificada como Merlus gayi. Kaup (1858) la incorporó al género Merluccius.
Debido a la imprecisión de los límites geográficos, esta especie fue confundida con M. hubbsi Marini, 1933 de Argentina. Norman (1937) restringió su distribución conocida a las costas de Chile y Perú, y llegó a confundirse con M. angustimanus Garman, 1899 de Panamá. Hildebrand (1946) describió la especie en detalle y discutió sus diferencias con M. productus (Ayres 1855) del Pacífico Norte, M. hubbsi y M. angustimanus (revisión de Inada 1981).
Reconocimiento de Subespecies y su Distribución Geográfica
Ginsburg (1954) reconoció dos subespecies en Merluccius gayi: M. gayi gayi (Guichenot 1848) y M. gayi peruanus Ginsburg, 1954, que habitan aguas de Chile y Perú, respectivamente (Leible 1979). Leible (1979) analizó la morfometría de ambos taxa y no encontró diferencias significativas para la clasificación subespecífica, pero sugirió mantener la nomenclatura trinomial debido a la separación geográfica.
Sin embargo, Inada (1981) encontró diferencias significativas en dos caracteres merísticos: el número de vértebras y de branquiespinas. Las subespecies (M. g. gayi y M. g. peruanus) presentan una distribución alopátrica, con una separación geográfica superior a los 1.300 km (Leible 1979), que se superpone con la cuenca de Chile-Perú. Las últimas revisiones estiman una separación mínima de 1.000 km, ya que el taxón del Perú (M. g. peruanus) se distribuye entre los 6ºS y los 14ºS (Espino et al. 1995) y el taxón de Chile (M. g. gayi) entre los 23º 39'S y los 47º 0'S (Aguayo 1995).
Hipótesis sobre el Origen de Merluccius gayi
Las hipótesis biogeográficas que explican el origen de M. gayi han sido planteadas por varios autores como Szidat (1955), Inada (1981), Kabata & Ho (1981), Fernández (1985), Ho (1990) y Stepien & Rosenblatt (1996). Exceptuando a Szidat (1955), quien plantea un origen desde el Pacífico, estos autores proponen que la especie se habría originado de un "stock" ancestral del Atlántico que migró hacia el Pacífico a través del Istmo de Panamá.
Kabata & Ho (1981) y Ho (1990), basándose en información parasitológica, sugieren que este paso se habría dado en el Oligoceno (24 a 38 millones de años atrás). Por su parte, Inada (1981) utilizando información osteológica, y Stepien & Rosenblatt (1996) usando información genética, postulan el Plioceno (2 a 6 millones de años). Sin embargo, hasta el momento no se había propuesto una hipótesis que intentara explicar el origen de las subespecies específicas de M. gayi.
Análisis de la Diferenciación Genética entre Subespecies
Problema y Enfoque del Estudio
Históricamente, los estudios sobre M. gayi se han centrado en caracteres merísticos y morfométricos, que son expresiones fenotípicas susceptibles de ser modificadas por la interacción genotipo-ambiente (Lewontin 1984). Esto dificulta determinar si las diferencias observadas son producto del ambiente y el tiempo actual (escala ecológica) o de la historia evolutiva y las presiones selectivas pasadas (escala evolutiva). Para resolver estos problemas sistemáticos, los estudios que utilizan información genética y parasitológica son cruciales.
Galleguillos et al. (1999) y Oyarzún (1997) han determinado la homogeneidad genética poblacional para M. g. gayi en las costas de Chile. En el presente trabajo, se analiza la diferenciación genética de ambos taxa (M. g. gayi y M. g. peruanus), se estima el tiempo de separación de las subespecies y se discuten los resultados considerando los antecedentes paleogeográficos del área de distribución.
Materiales y Métodos Utilizados
Para analizar la diferenciación genética de los taxa, se tomó una muestra de 25 individuos de M. g. gayi de la zona de Talcahuano (Chile) (36º 42' 14" S; 73º 05' W) y se recolectaron 25 individuos de M. g. peruanus del Puerto de Paita en Perú (5º 05' S; 81º 10' W). Las merluzas se mantuvieron a -20º C y a cada individuo se le extrajo un trozo de músculo esquelético.
El músculo se homogeneizó y centrifugó, y el sobrenadante se absorbió con papel filtro para ser montado en un gel de almidón al 12% y aplicar la técnica de electroforesis horizontal. Las proteínas fueron teñidas siguiendo las técnicas histoquímicas de Harris & Hopkinson (1976) y Ward & Galleguillos (1983). Se determinó el tampón con mejor resolución, los loci de cada taxón y el carácter monomórfico o polimórfico de cada locus. La nomenclatura de las enzimas siguió las sugerencias de Shaklee et al. (1990). Los alelos de cada locus se denominaron según su aparición en el gel desde el origen (A, B y C). Se consideraron polimórficos aquellos loci cuya frecuencia del alelo más común fue menor o igual a 0,95. Finalmente, se compararon los taxa en cuanto a su heterocigosidad y polimorfismo, y se calcularon los índices estándar de Identidad (I) y Distancia genética (D) (Nei 1978) utilizando el programa BIOSYS 1.7 (Swofford & Selander 1989). A partir de estos índices, se calculó el tiempo de separación de acuerdo a la hipótesis del reloj molecular (Nei 1978), usando estimaciones de Vawter et al. (1980) y Smith & Coss (1984).
Resultados de la Diferenciación Genética
Para la subespecie M. g. gayi, se determinaron 27 loci enzimáticos y 5 loci de proteínas estructurales. Se observaron 5 loci enzimáticos polimórficos (MDH-2*, PGM-1*, EST-3*, CA-2*, GPI-1*), de los cuales el locus GPI-1* presentó la mayor heterocigosidad esperada según Hardy-Weinberg (0,49). Solo el locus PT-4* no se ajustó a las frecuencias esperadas (P = 0,04). Esta subespecie mostró la mayor variabilidad genética, con un polimorfismo del 24% y una media de heterocigosidad esperada de 0,072 ± 0,018.
En M. g. peruanus, se analizaron 6 loci de proteínas totales y 29 loci enzimáticos, de los cuales 3 resultaron polimórficos (PGM-1*, EST-3*, GPI-1*). Todos los loci de este taxón se ajustaron al equilibrio de Hardy-Weinberg (P > 0,1). El valor de heterocigosidad esperado más alto se determinó para el locus GPI-1* (0,44). Respecto a la variabilidad genética, esta subespecie presentó un polimorfismo del 18% y una media de heterocigosidad esperada de 0,037 ± 0,018, valores más bajos que los encontrados para M. g. gayi.
De los loci observados, 3 monomórficos (EM-2*, AP-1* y CA-1*) y uno polimórfico (EST-3*) no presentaron alelos compartidos por los dos taxa, lo que los constituye en marcadores genéticos de tipo diagnóstico. Ambos taxa presentaron alelos únicos para otros marcadores, aunque con baja frecuencia. Destaca que los loci EST-1*, AP-3* y PT-0* solo se observaron en M. g. peruanus, sirviendo como caracteres diagnósticos para este taxón. Las diferencias en variabilidad genética y migración de alelos permitieron determinar una identidad genética (I) de 0,873 y una distancia genética (D) de 0,136. Esto, llevado a tiempo de separación según la hipótesis del reloj molecular, corresponde a una divergencia evolutiva de 900.000 a 2.000.000 de años entre M. g. gayi y M. g. peruanus.
Discusión y Evidencia Paleogeográfica
Los valores relativamente bajos de heterocigosidad observados en este trabajo coinciden con los obtenidos en otras especies de merluzas (Shaklee et al. 1982). M. g. peruanus posee una heterocigosidad muy similar a la de M. productus (0,041 ± 0,012) (Stepien & Rosenblatt 1996), pero más baja que la de M. australis (0,044 ± 0,016) (Hernández 1996), M. capensis (0,051), M. paradoxus (0,063) (Grant et al. 1988), M. hubbsi (0,066 ± 0,02) y una muestra de M. g. gayi (0,051 ± 0,015) del Pacífico Sureste (Stepien & Rosenblatt 1996).
Por otro lado, la muestra de M. g. gayi analizada en este estudio presentó valores de heterocigosidad y polimorfismo mayores que todas las especies citadas. La homogeneidad genética de M. gayi en la costa de Chile ha sido verificada por Galleguillos et al. (1999) y reafirmada por análisis de ADN mitocondrial (Oyarzún 1997). La variabilidad morfológica también ha sido analizada, mostrando patrones propios de una especie de amplia distribución latitudinal (Oyarzún 1997), sin formar grupos discretos.
La distribución alopátrica de los grupos geográficos, la información genético-poblacional, el número de branquiespinas y el número promedio de sustituciones alélicas (resultado de la evolución independiente de los taxa) (Nei 1978, Ayala & Kiger 1980, Buth 1984), permiten confirmar la categoría al menos subespecífica para las merluzas Chilena y Peruana. Los valores de distancia genética obtenidos para estas subespecies son similares a los encontrados para el reconocimiento subespecífico en la familia Pleuronectidae (Ward & Galleguillos 1983) y en el género Lepomis (Avise & Smith 1977). La existencia de tres loci únicos para M. g. peruanus reafirma su condición subespecífica, concordando con la propuesta original de Ginsburg (1954).
El valor de distancia genética encontrado es muy similar al informado por Stepien & Rosenblatt (1996) para la comparación entre M. gayi y M. productus (D = 0,132), pero menor al comparar M. gayi con M. hubbsi (D = 0,276) y M. hubbsi con M. productus (D = 0,367), y menor que los informados por Hernández (1996) entre M. australis y M. g. gayi (D = 0,430) y M. australis con M. g. peruanus (D = 0,330). Esta situación sugiere que los taxa de merluzas del Pacífico Oriental se encuentran en una etapa avanzada de especiación y que su estatus taxonómico corresponde al nivel subespecífico, aunque se requieren otros estudios (como la comparación del ADN mitocondrial) para sustentar o rechazar un nivel taxonómico específico.
Según Inada (1981), Kabata & Ho (1981), Fernández (1985), Ho (1990) y Stepien & Rosenblatt (1996), los taxa de merluzas del Pacífico Oriental se habrían originado de una población ancestral que migró inicialmente hacia el Pacífico a través del Istmo de Panamá, el cual se cerró en el Plioceno, hace 3 millones de años (Kennett 1982). El nivel de diferenciación genética encontrado para las subespecies de M. gayi, que equivale a una separación de entre 900.000 y 2.000.000 de años, permitiría establecer que la población ancestral estuvo inicialmente ligada a la plataforma continental del norte del Perú. Su migración hacia el sur estuvo inicialmente restringida por la gran cuenca Chile-Perú, un evento geográfico formado antes del Mesozoico.
Esta barrera geográfica restringiría la plataforma continental, zona a la cual se asocian las especies de merluzas por hábitos alimenticios y reproductivos (Stepien & Rosenblatt 1996). De acuerdo a los tiempos de separación estimados, esta barrera pudo haber sido flanqueada durante alguno de los períodos interglaciales en que subió el nivel del mar durante el Pleistoceno. Esta hipótesis es respaldada por la información paleogeográfica, ya que, según Cecioni (1970), durante el Pleistoceno el mar avanzó en áreas de las costas del norte de Chile y el sur del Perú, lo que habría permitido la formación de bahías y la extensión de la plataforma continental sumergida hacia tierras antes emergidas.
LA EXTINCIÓN DE LA MEGA FAUNA DE LA EDAD DE HIELO - DOCUMENTAL
Descripción Física de Merluccius gayi
Merluccius gayi (Guichenot, 1848), conocida como merluza sureña o merluza del Pacífico sur, posee un cuerpo alargado, comprimido, y se afina hacia una delgada base de la cola. Su cabeza es larga (26-32% de la longitud estándar, LE) y achatada, con una cresta en forma de "V" en el dorso. Los ojos son grandes y la boca también es grande, con la mandíbula inferior ligeramente proyectada. Los dientes de las mandíbulas son largos, fuertes y puntiagudos, dispuestos en dos filas irregulares. Presenta dientes en el centro del paladar, pero no en los costados.
- Branquiespinas: 18-25
- Aletas dorsales: Dos aletas dorsales separadas; la primera es más corta y triangular (I, 9-12 radios), la segunda tiene una base larga y está parcialmente dividida por una escotadura (36-42 radios).
- Aleta anal: Similar a la segunda dorsal, con 36-42 radios.
- Aletas pélvicas: Bien desarrolladas y no filamentosas, con 7 radios, situadas delante de las aletas pectorales.
- Aleta caudal: Corta y ligeramente ahorquillada.
- Escamas: Pequeñas, con 106-130 en la línea lateral.
El color del dorso es azul metálico, y los flancos son plateados. Puede alcanzar un tamaño de 87 cm. Su hábitat es pelágico, y se encuentra a profundidades de 50-500 metros. Su distribución abarca desde Chile hasta Ecuador, incluyendo las Islas Galápagos. M. gayi incluye dos subespecies: M. g. peruanus, que se encuentra en Ecuador y Perú, y M. g. gayi, que habita las costas de Chile.